Navigate toolbar: [ Diets | Photos | Estuarine | FADs | Soniferous | CV | Home Page ]
Analysis for temporal trophic structure changes paper
Fogarty, M.J., M. Grosslein and R.A. Rountree. Structural changes in an exploited marine system. Nature
Summary
: Examines community and trophic structure changes on Georges Bank during a period of dramatic population declines of commercial species (1973-90). Both population abundance and trophic data sets reveal that the George Bank fish community has shifted from a Gadid and flatfish dominated demersal fish community to one dominated by non-commercial pelagic/semi-pelagic (mackerel, herring, and spiny dogfish) and demersal species (skates and sculpins).Note: this paper is very far on the back burner at this time due to the difficulty of using NMFS data to examine temporal changes in trophic structure.
The trophic analysis for this paper has been particularly difficult due to inherent problems with the food habitats time series. Changes in sampling priorities and methods used by the National Marine Fisheries Service in their food habits data collections make it very difficult to analyze time-series changes in Georges Bank (or other geographic areas) trophic structure. Sufficient data was often not collected for any given species over the entire 18 year time series (1973-90),much less for a consistent suite of fishes. This problem is obvious even when data is pooled overall size classes and geographic areas:
Table 1. Number of stomach samples by predator species and year pooled over all seasons, geographic areas and predator size classes.
Table 2. Number of stomach samples by predator species, season and year pooled over all geographic areas and predator size classes.
Table 3. Number of stomach samples by predator species, year and geographic area pooled over seasons.
To get around the weaknesses in the time series I have attempted to examine temporal trends in trophic structure in several different ways, all of which have various strengths and weaknesses: I. group years and predator size classes to increase sample sizes among treatment group, II. Conduct separate analyzes for each treatment group and compare predator-prey relationships, III. Conduct time series analyzes for each predator to look for changes in its feeding habitats over time, compare species trends to look for overall trends.
In the first method, predators were lumped into one or two size classes and data were grouped into two time periods (1981-85 and 1986-90) and two seasons (spring and fall), for Georges Bank data only. Canonical discrimination analysis (CDA) was then used to directly test for differences in the trophic structure among time period and season groups. Cluster analysis and PCA analysis were also performed, but proved to variable and subjective to present. CDA, and Correspondance analysis (CA, see below), are also subjective in terms of identifying trophic guilds, but not in terms of the output. In contrast, many dendrograms can be produced with cluster analysis on the same data.
Table 4. Predator species, and total sample sizes for each period and season used in the analysis.
Table 5. Sample sizes for each predator by time period, season, and length group.
Table 6. Predator by prey matrices of standardized weighted mean percent stomach volumes for each period and season (raw data going into analyzes).
Table 7. SAS output from the CDA analysis.
Figure 1. Graph of cluster of each predator species in canonical space with 95 CI drawn around the group means. The group effect was highly significant (see statistics in Table 7). There is an interaction effect between the two canonicals, so that both are needed to separate out the groups. Prey with high loadings on the canonicals are shown by the vectors drawn from the origin. In addition silver hake, Urophycis sp., L.h. sculpin, and fawn cusk eel prey were significantly positively correlated with CAN2, D. leptocerus, Gadidae, and Scombridae were negatively correlated with Can2, haddock was negatively, and pandalidae was positively correlated with Can1. This analysis clearly suggests strong changes in trophic structure between early and late 1980s,including decreasing separation between spring and fall. The weakness of this analysis is that because of grossly unequal sampling among size classes and years we can't be sure that the group differences are real, or result from the sampling bias. I have already shown elsewhere (see feeding types paper), that even small differences in the distribution in size classes over which data are pooled can led to large differences in the pooled diet estimation. So, for example, the greaterimportance of herring in the 1986-90 period relative to the 1981-85 period could result from a greater proportion of larger predators in the later period.
Figure 2. CDA analysis testing for differences among years from 1977-1988 based on diets of 11species (unweighted mean stomach volumes/weights per trawl tow). In this analysis prey were grouped to the highest taxa (phyla or pseudophyla) to allow comparison between the 1970's and1980's where different methods were used. There was a significant group effect where the 70'syears cluster together and far from the 80's years. The 80's cluster into two groups: 1981-82, and1983-88. A MANOVA suggests that the 70's were highly significantly different from the 80's.This pattern seems to be driven mostly by spiny dogfish, winter skate , silver hake and white hake, in that order. But note there is also a lot of differences among years in the 80's, suggesting that arbitrarily pooling over years, as was done above, may be problematic.
Table 8. Diet similarity matrices among predators for each of the 4 time period and season groups (81-85, 86-90, spring, fall).
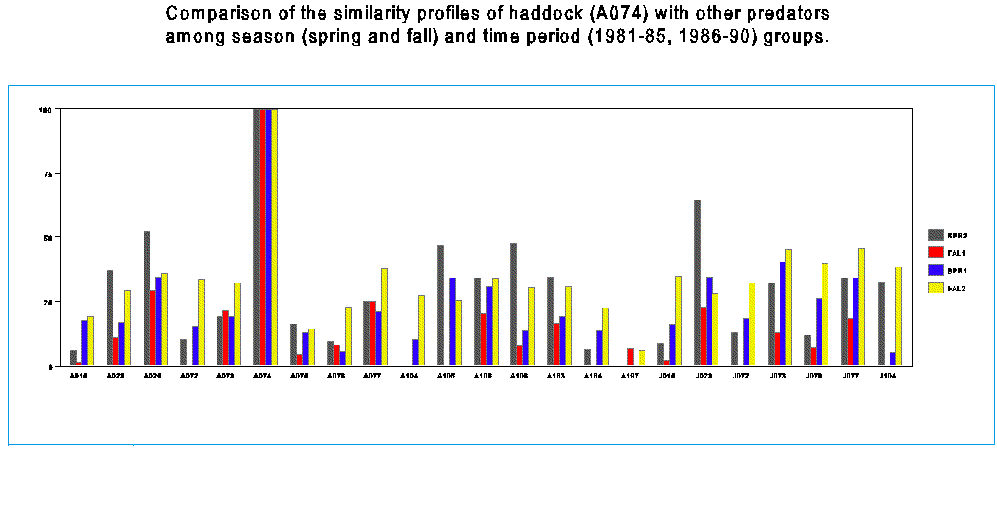
Table 9. Same as Table 8, Diet similarity matrix among predators and treatment groups resorted to allow comparison among treatment groups for each predator. Each line of this table can be viewed as a profile, or vector of the diet similarity of a given predator to all the other predators under consideration. I reasoned that an examination of changes in this profile among the treatment groups could be used to infer changes in the trophic structure.
Figure 2a. For example, a comparison of the similarity profiles of haddock to other predators among the four treatment group suggests strong seasonal and time period changes in the potential for competition between haddock and other Georges bank groundfishes.
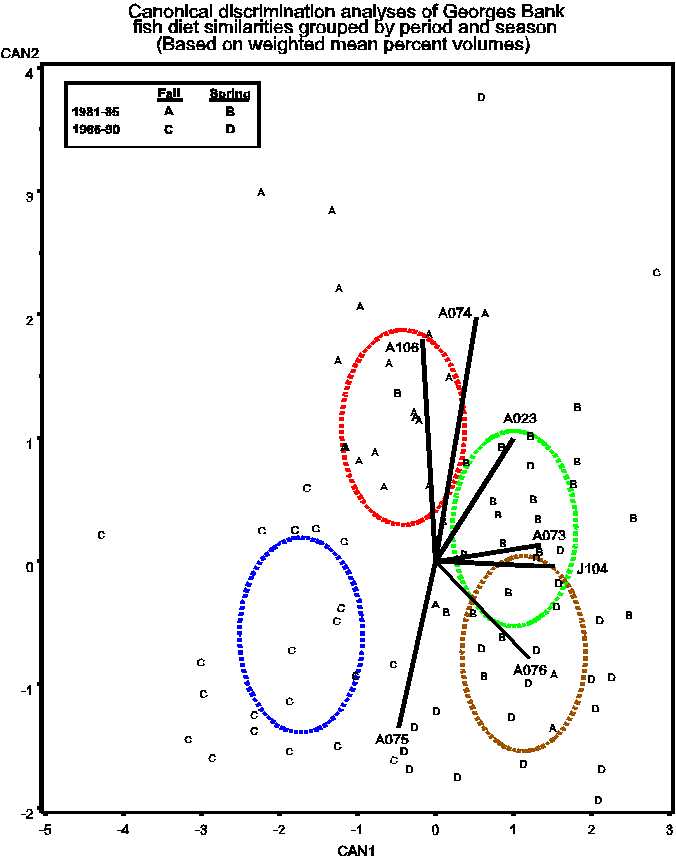
Figure 3. CDA analysis based on the similarity matrix (actually used the dissimilarity matrix for the computations) with four treatment groups. This analysis was highly significant (78), Can1(59) and Can2 (36 ) appear to separate the groups among seasons and time periods, respectively. Large haddock (A074), large winter flounder (A106), and large pollock (A075)have the strongest influence on time period separation (can2). I interpret this as indicating that the similarities of these species to all other species changes between time periods, which indeed can be seen in Table 9 and Figure 2a. Problem is I'm not sure that applying CDA analysis to a Pred*pred similarity matrix is statistically valid.
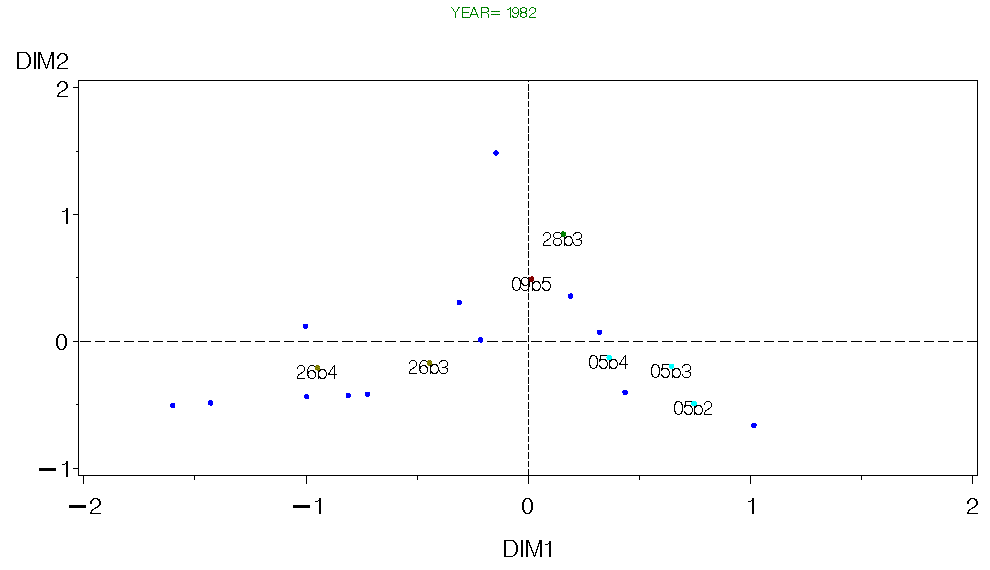
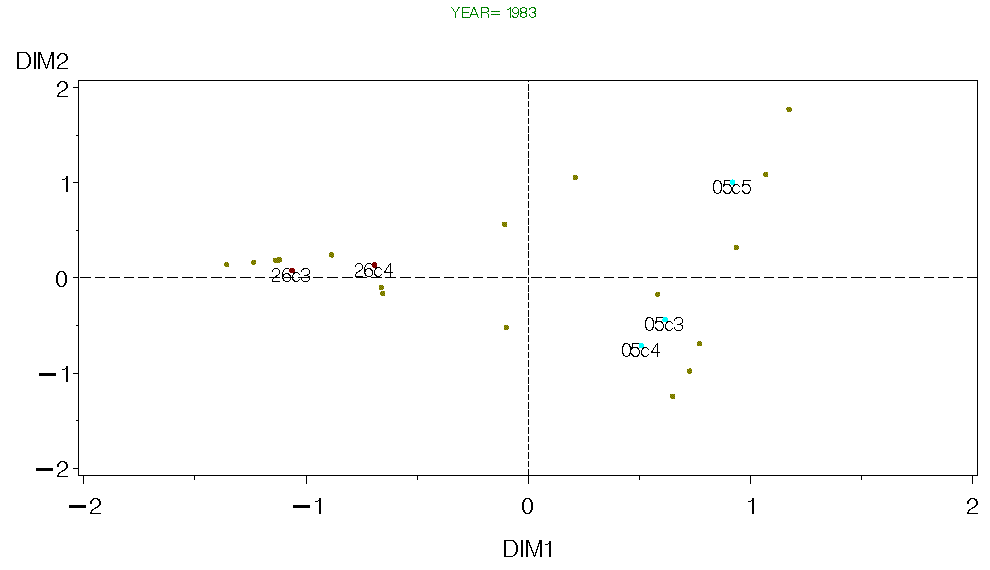
A second approach is to avoid pooling data by conducting separate analysis for each year and season in the time series. Each predator species is divided up into up to 5 size classes based on a percentage of the predator's maximum size (consistent with methods used in guild and feeding types analyzes). All size classes with at least 20 samples for a given year and season were included in the analyzes. All of the above analyzes were severely hampered by the need to have all predator species/size classes in each treatment group. The only way to do that was to pool over many years/seasons and size classes. Even so, we were left with only a few species to include in our trophic system. Alternatively, if we conduct separate analysis of the trophic structure for each year we can avoid pooling to a large degree. We can then subjectively compare the results among years. The problem here, of course, is that since we essentially have different suites of predators and prey during each year, we are in danger of comparing apples and oranges among years. However, this approach could reveal general patterns, and possibly changes in these patterns through time. I used correspondance analysis (CA) to examine the relationships among predators and prey for each year and season. I show results from Fall 1982-90 below. The intention is to define guild structures for each year and seasons on Georges bank as was done for each geographic area using pooled data (see Guild page). One big problem with this approach is that we have data on two few species to adequately define species "guilds". However, for species common to each data set, there does appear to be some similar trends, so it may be possible to infer changes in trophic structure with this approach.
(Note, these are quick and dirty raw figures, if I pursue this approach figures similar to those used in the guild analysis would be made. For some semblance of clarity, the prey scores are not labeled, but their distributions are plotted with the colored dots).
Figure 4. CA for Georges Bank Fall 1982.
Figure 5. CA for Georges Bank Fall 1983.
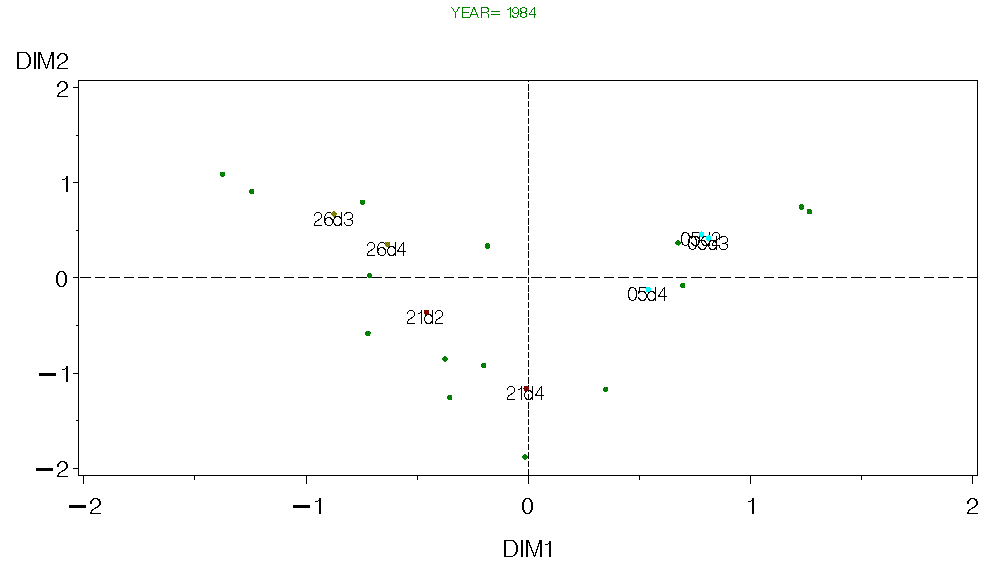
Figure 6. CA for Georges Bank Fall 1984.
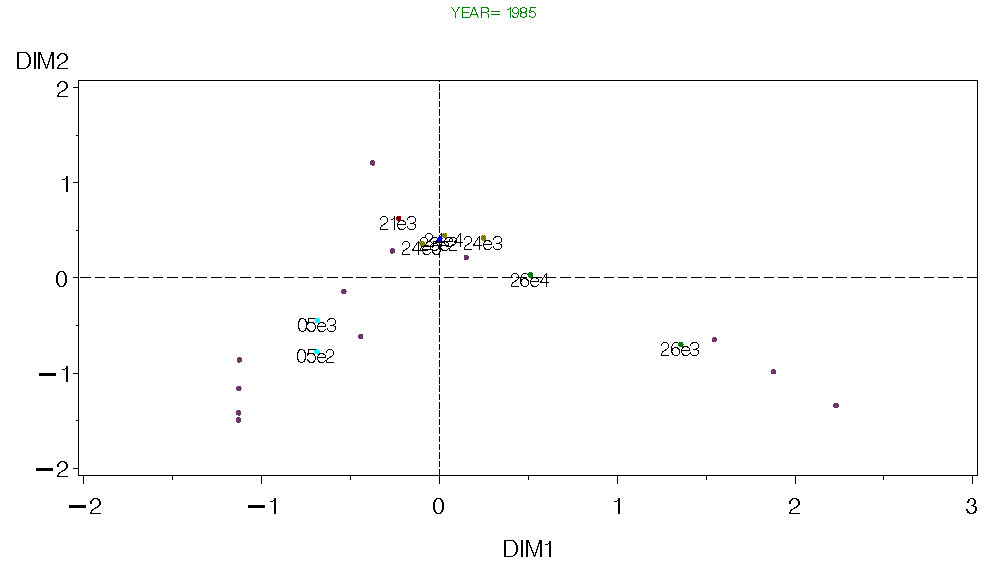
Figure 7. CA for Georges Bank Fall 1985.
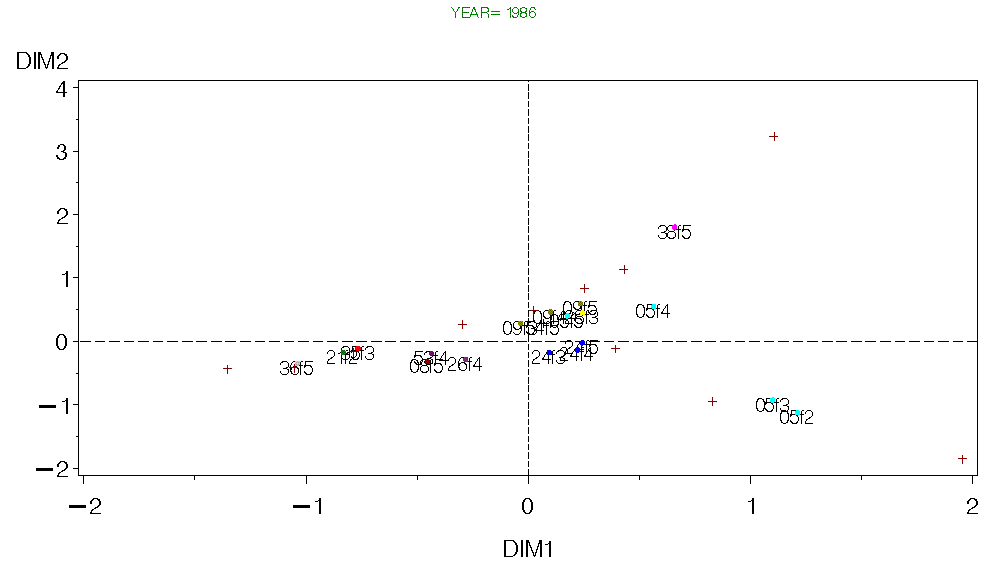
Figure 8. CA for Georges Bank Fall 1986.
Figure 9. CA for Georges Bank Fall 1987.
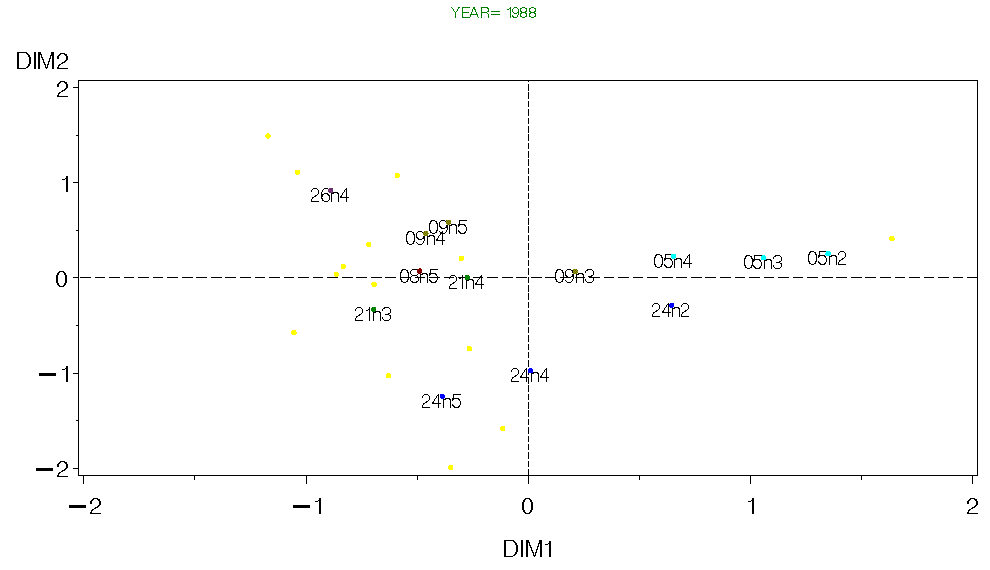
Figure 10. CA for Georges Bank Fall 1988.
Figure 11. CA for Georges Bank Fall 1989.
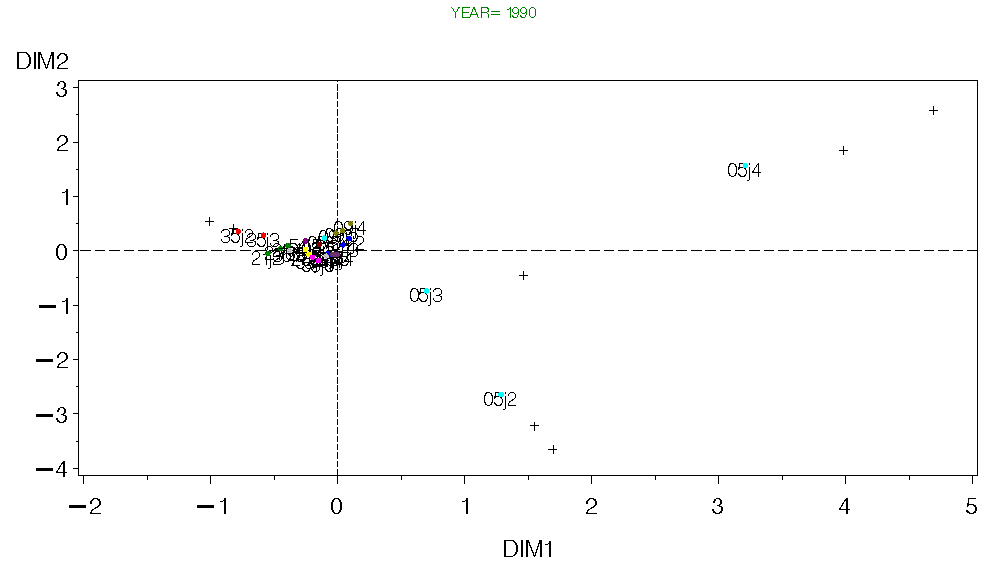
Figure 12. CA for Georges Bank Fall 1990.
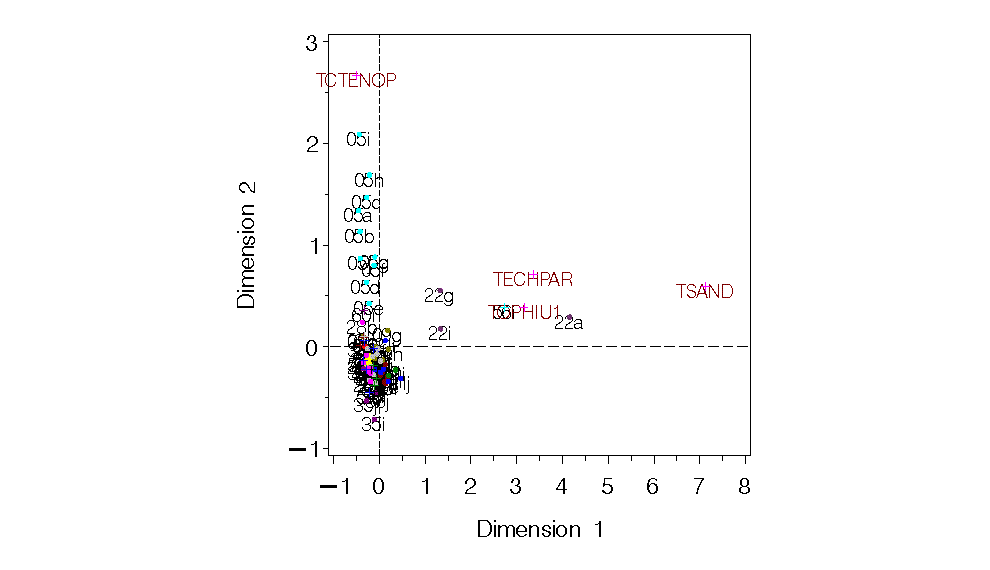
Figure 13. CA for Georges Bank all species and years combined in single analysis. No obvious linear temporal trend.
The third method is to not attempt to examine species interrelationships directly, but to examine temporal trends in each predator's diet separately. A comparison of trends among species can then be made to infer temporal changes in trophic structure. I've done a bunch of exploratory analysis here. For example, I've done both CDA and CD analysis for all the species to look at differences among years (just quick and dirty, too much to print out). Eyeballing the results does suggest that some species may exhibit linear trends in diet (e.g. spiny dogfish), but most species exhibit no logical temporal trends. Often lots of differences between years, but no logical pattern.
Well at least you know what I've been doing. In some ways the approach used in Method I, is the best. I still like Figures 1 and 3 a lot. Method II is more like Mike suggested some time ago using cluster analysis. First species "guilds" and prey defining them, are determined for each year and then subjectively compared among years. Only problem here, is its tedious to make all the figures for each year and season, and then how do you define a "guild" from the CA plots (see guild analysis). Method III is appealing in that the sample bias problems are eliminated. Each species might have a different time series, with various holes in the time series, but that's O.K. Here we would not be directly examining trophic structure, but could infer basic changes in the community based on similar patterns among species. Let me know what you think.
This page was last modified on July 20, 2001
Copyright © 1999 by Rodney Rountree. All rights reserved
Navigate to main diet pages: [ Parent | Summary | Fluke | GMDiet | GMGuild | Goosefish | Guild | Ontogenetic | Prey size | Structure ]
Navigate to main pages: [ Diets | Photos | Estuarine | FADs | Soniferous | CV | Home Page ]
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}